Физиология боли
В узком смысле слова боль – это неприятное ощущение, возникающее при действии сверхсильных раздражителей, вызывающих структурно-функциональные нарушения в организме. Отличия боли от других ощущений в том, что она не информирует мозг о качестве раздражителя, а указывает на то, что раздражитель является повреждающим. Другой особенностью болевой сенсорной системы является наиболее сложной и мощный ее эфферентный контроль.
Болевой анализатор запускает в ЦНС несколько программ ответа организма на боль. Следовательно, боль имеет несколько компонентов. Сенсорный компонент боли характеризует ее как неприятное, тягостное ощущение; аффективный компонент – как сильную отрицательную эмоцию; мотивационный компонент – как отрицательную биологическую потребность, запускающую поведение организма, направленное на выздоровление. Моторный компонент боли представлен различными двигательными реакциями: от безусловных сгибательных рефлексов до двигательных программ антиболевого поведения. Вегетативный компонент характеризует нарушение функций внутренних органов и обмена веществ при хронических болях. Когнитивный компонент связан с самооценкой боли, боль при этом выступает как страдание. При деятельности других систем эти компоненты слабо выражены.
Биологическая роль боли определяется несколькими факторами. Боль исполняет роль сигнала об угрозе или повреждении тканей организма и предупреждает их. Боль имеет познавательную функцию: человек через боль учится избегать возможных опасностей внешней среды. Эмоциональный компонент боли выполняет функцию подкрепления при образовании условных рефлексов. Боль является фактором мобилизации защитно-приспособительных реакций организма при повреждении его тканей и органов.
Выделяют два вида боли – соматическую и висцеральную. Соматическую боль подразделяют на поверхностную и глубокую Поверхностная боль может быть ранняя (быстрая, эпикрическая) и поздняя (медленная, протопатическая).
Существуют три теории боли.
1. Теория интенсивности была предложена Э.Дарвином и А.Гольдштейнером. По этой теории боль не является специфическим чувством и не имеет своих специальных рецепторов. Она возникает при действии сверхсильных раздражителей на рецепторы пяти известных органов чувств. В формировании боли участвуют конвергенция и суммация импульсов в спинном и головном мозге.
2. Теория специфичности была сформулирована немецким физиологом М.Фреем. В соответствии с этой теорией боль является специфическим чувством, имеющим собственный рецепторный аппарат, афферентные волокна и структуры головного мозга, перерабатывающие болевую информацию. Эта теория в дальнейшем получила более полное экспериментальное и клиническое подтверждение.
3. Современная теория боли базируется преимущественно на теории специфичности. Было доказано существование специфичных болевых рецепторов. Вместе с тем в современной теории боли использовано положение о роли центральной суммации и конвергенции в механизмах боли. Наиболее крупными достижениями современной теории боли является разработка механизмов центрального восприятия боли и запуска противоболевой системы организма.
Болевые рецепторы
Болевые рецепторы являются свободными окончаниями чувствительных миелиновых нервных волокон Аδ и немиелиновых волокон С. Они найдены в коже, слизистых оболочках, надкостнице, зубах, мышцах, суставах, внутренних органах и их оболочках, сосудах. Их нет в нервной ткани головного и спинного мозга. Наибольшая их плотность имеется на границе дентина и эмали зуба.
Выделяют следующие основные типы болевых рецепторов:
1. Механоноцицепторы и механотермические ноцицепторы Аδ-волокон реагируют на сильные механические и термические раздражители, проводят быструю механическую и термическую боль, быстро адаптируются; расположены преимущественно в коже, мышцах, суставах, надкостнице; их афферентные нейроны имеют малые рецептивные поля.
2. Полисенсорные ноцицепторы С-волокон реагируют на механические, термические и химические раздражители, проводят позднюю плохо локализованную боль, медленно адаптируются; их афферентные нейроны имеют большие рецептивные поля.
Болевые рецепторы возбуждаются тремя видами раздражителей:
1. Механические раздражители, создающие давление более 40г/мм 2 при сдавливании, растяжении, сгибании, скручивании.
2. Термические раздражители могут быть тепловыми (> 45 0 С) и холодовыми (< 15 0 С).
3. Химические раздражители, освобождающиеся из поврежденных клеток тканей, тучных клеток, тромбоцитов (К + , Н + , серотонин, ацетилхолин, гистамин), плазмы крови (брадикинин, каллидин) и окончаний ноцицептивных нейронов (вещество Р). Одни из них возбуждают ноцицепторы (К + , серотонин, гистамин, брадикинин, АДФ), другие сенсибилизируют их.
Свойства болевых рецепторов: болевые рецепторы имеют высокий порог возбуждения, что обеспечивает их ответ только на чрезвычайные раздражители. Ноцицепторы С-афферентов плохо адаптируются к длительно действующим раздражителям. Возможно повышение чувствительности болевых рецепторов – снижение порога их раздражения при многократной или длительной стимуляции, что называется гипералгезией. При этом ноцицепторы способны отвечать на стимулы субпороговой величины, а также возбуждаться раздражителями других модальностей.
Проводящие пути болевой чувствительности
Нейроны, воспринимающие болевую импульсацию. От болевых рецепторов туловища, шеи и конечностей Аδ- и С-волокна первых чувствительных нейронов (их тела находятся в спинальных ганглиях) идут в составе спинномозговых нервов и входят через задние корешки в спинной мозг, где разветвляются в задних столбах и образуют синаптические связи прямо или через интернейроны со вторыми чувствительными нейронами, длинные аксоны которых входят в состав спиноталамических путей. При этом они возбуждают два вида нейронов: одни нейроны активируются только болевыми стимулами, другие – конвергентные нейроны – возбуждаются также и неболевыми стимулами. Вторые нейроны болевой чувствительности преимущественно входят в состав боковых спиноталамических путей, которые и проводят большую часть болевых импульсов. На уровне спинного мозга аксоны этих нейронов переходят на сторону, противоположную раздражению, в стволе головного мозга они доходят до таламуса и образуют синапсы на нейронах его ядер. Часть болевой импульсации первых афферентных нейронов переключаются через интернейроны на мотонейроны мышц-сгибателей и участвуют в формировании защитных болевых рефлексов. В боковом спиноталамическом пути выделяют эволюционно более молодой неоспиноталамический путь и древний палеоспиноталамический путь.
Неоспиноталамический путь проводит болевые сигналы по Аδ-волокнам преимущественно в специфические сенсорные (вентральные задние) ядра таламуса, имеющие хорошую топографическую проекцию периферии тела. Кроме этого небольшая часть импульсов поступает в ретикулярную формацию ствола и далее в неспецифические ядра таламуса. Передача возбуждения в синапсах этого пути осуществляется с помощью быстродействующего медиатора глутамата. Из специфических ядер таламуса болевые сигналы передаются преимущественно в сенсорную кору больших полушарий. Эти особенности формируют основную функцию неоспиноталамического пути – проведение «быстрой» боли и восприятие ее с высокой степенью локализации.
Палеоспиноталамический путь проводит болевые сигналы по С-волокнам преимущественно в неспецифические ядра таламуса прямо или после переключения в нейронах ретикулярной формации ствола мозга. Передача возбуждения в синапсах этого пути происходит более медленно. Медиатором является вещество Р. Из неспецифических ядер импульсация поступает в сенсорную и другие отделы коры больших полушарий. Небольшая часть импульсации поступает и в специфические ядра таламуса. В основном волокна этого пути оканчиваются на нейронах 1) неспецифических ядер таламуса; 2) ретикулярной формации; 3) центрального серого вещества; 4) голубого пятна; 5) гипоталамуса. Через палеоспиноталамический путь проводится «поздняя», плохо локализуемая боль, формируются аффективно-мотивационные проявления болевой чувствительности.
Кроме этого болевая чувствительность частично проводится по другим восходящим путям: переднему спиноталамическому, тонкому и клиновидному путям.
Вышеназванные пути проводят и другие виды чувствительности: температурную и тактильную.
Роль коры больших полушарий в восприятии боли
Полноценное чувственное восприятие боли организмом без участия коры головного мозга невозможно.
Первичное проекционное поле болевого анализатора находится в соматосенсорной коре задней центральной извилины. Оно обеспечивает восприятие «быстрой» боли и идентификацию места ее возникновения на теле. Для более точной идентификации локализации боли в процесс обязательно включается и нейроны моторной коры передней центральной извилины.
Вторичное проекционное поле расположено в соматосенсорной коре на границе пересечения центральной борозды с верхним краем височной доли. Нейроны данного поля имеют двусторонние связи с ядрами таламуса, что позволяет этому полю избирательно фильтровать, проходящие через таламус возбуждения болевого характера. А это в свою очередь позволяет данному полю вовлекаться в процессы, связанные с извлечением из памяти энграммы необходимого поведенческого акта, его реализации в деятельности эффекторов и оценки качества достигнутого полезного результата. Двигательные компоненты болевого поведения формируются в совместной деятельности моторной и премоторной коры, базальных ганглиев и мозжечка.
Лобная кора играет важную роль в восприятии боли. Она обеспечивает самооценку боли (ее когнитивный компонент) и формирование целенаправленного болевого поведения.
Лимбическая система (поясная извилина, гиппокамп, зубчатая извилина, миндалевидный комплекс височной доли) получает болевую информацию от передних ядер таламуса и формирует эмоциональный компонент боли, запускает вегетативные, соматические и поведенческие реакции, обеспечивающие приспособительные реакции к болевому раздражителю.
Некоторые виды болевых ощущений
Существуют боли, которые названы проекционными или фантомными . Их возникновение основано на законе проекции боли: какая бы часть афферентного пути не раздражалась, боль ощущается в области рецепторов данного сенсорного пути. По современным данным в формировании данного вида болевого ощущения участвуют все отделы болевой сенсорной системы.
Существуют также так называемые отраженные боли: когда боль ощущается не только в пораженном органе, но и в соответствующем дерматоме тела. Участки поверхности тела соответствующего дерматома, где возникает ощущение боли, назвали зонами Захарьина – Геда . Возникновение отраженных болей связано с тем, что нейроны, проводящие болевую импульсацию от рецепторов пораженного органа и кожи соответствующего дерматома, конвергируют на одном и том же нейроне спиноталамического пути. Раздражение этого нейрона с рецепторов пораженного органа в соответствии с законом проекции боли приводит к тому, что боль ощущается и в области кожных рецепторов.
Антиноцицептивная система
Антиболевая система состоит из четырех уровней: спинального, стволового, гипоталамического и коркового.
1. Спинальный уровень антиноцицептовной системы. Важным ее компонентом является «воротный контроль» спинного мозга, концепция которого имеет следующие основные положения: передача болевых нервных импульсов с первых нейронов на нейроны спиноталамических путей (вторые нейроны) в задних столбах спинного мозга модулируется спинальным воротным механизмом – тормозными нейронами, расположенными в желатинозном веществе спинного мозга. На этих нейронах оканчиваются разветвления аксонов различных сенсорных путей. В свою очередь нейроны желатинозной субстанции оказывают пресинаптическое торможение в местах переключения первых и вторых нейронов болевых и других сенсорных путей. Часть нейронов являются конвергентными: на них образуют синапсы нейроны не только от болевых, но и от других рецепторов. Спинальный воротный контроль регулируется соотношением импульсов, поступающих по афферентным волокнам большого диаметра (неболевая чувствительность) и малого диаметра (болевая чувствительность). Интенсивный поток импульсов по волокнам большого диаметра ограничивает передачу болевых сигналов на нейроны спиноталамических путей (закрывает «ворота»). Напротив, интенсивный поток болевых импульсов по первому афферентному нейрону, ингибируя тормозные интернейроны, облегчает передачу болевых сигналов на нейроны спиноталамических путей (открывает «ворота»). Спинальный воротный механизм находится под постоянным влиянием нервных импульсов структур ствола мозга, которые передаются по нисходящим путям как на нейроны желатинозной субстанции, так и на нейроны спиноталамических путей.
2. Стволой уровень антиноцицептивной системы. К стволовым структурам противоболевой системы относятся, во-первых, центральное серое вещество и ядра шва, образующие единый функциональный блок, во-вторых, крупноклеточное и парагигантоклеточное ядра ретикулярной формации и голубое пятно. Первый комплекс блокирует прохождение болевой импульсации на уровне релейных нейронов ядер задних рогов спинного мозга, а также релейных нейронов сенсорных ядер тройничного нерва, образующих восходящие пути болевой чувствительности. Второй комплекс возбуждает почти всю антиноцицептивную систему (см.рис.1).
3. Гипоталамический уровень антиноцицептивной системы, с одной стороны, функционирует самостоятельно, а с другой – выступает как настройка, контролирующая и регулирующая антиноцицептивные механизмы стволового уровня за счет связей гипоталамических нейронов разной ядерной принадлежности и разной нейрохимической специфичности. Среди них идентифицированы нейроны, в окончаниях аксонов которых выделяются энкефалины, β-эндорфин, норадреналин, дофамин см.рис.2).
4. Корковый уровень антиноцицептивной системы. Объединяет и контролирует деятельность антиноцицептивных структур различного уровня соматосенсорная область коры больших полушарий. При этом наиболее важную роль в активации спинальных и стволовых структур играет вторичная сенсорная область. Ее нейроны образуют наибольшее количество волокон нисходящего контроля болевой чувствительности, направляющиеся к задним рогам спинного мозга и ядрам ствола головного мозга. Вторичная сенсорная кора видоизменяет активность стволового комплекса антиноцицептивной системы. Кроме этого соматосенсорные поля коры больших полушарий контролируют проведение афферентных болевых импульсов через таламус. Кроме таламуса, кора большого мозга регламентирует прохождение болевой импульсации в гипоталамусе, лимбической системе, ретикулярной формации, спинном мозге. Ведущая роль в обеспечении кортико-гипоталамических влияний отводится нейронам лобной коры.
Медиаторы антиноцицептивной системы
К медиаторам противоболевой системы относят пептиды, которые образуются в головном мозге, аденогипофизе, мозговом слое надпочечников, желудочно-кишечном тракте, плаценте из неактивных предшественников.. Сейчас к опиатным медиаторам антиноцицептивной системы относят: 1) ά-, β-, γ-эндорфины; 2) энкефалины; 3) динорфины. Эти медиаторы действуют на три вида опиатных рецепторов: μ-, δ-, κ-рецепторы. Наиболее селективным стимулятором μ-рецепторов являются эндорфины, δ-рецепторов – энкефалины, а κ-рецепторов – динорфины. Плотность μ- и κ-рецепторов высокая в коре больших полушарий и в спинном мозгу, средняя – в стволе головного мозга; плотность δ-рецепторов средняя в коре больших полушарий и спинном мозгу, малая – в стволе мозга. Опиоидные пептиды угнетают действие веществ, вызывающих боль, на уровне ноцицепторов, уменьшают возбудимость и проводимость болевой импульсации, угнетают вызванную реакцию нейронов, находящихся в составе цепей, передающих болевую импульсацию. Эти пептиды поступают к нейронам болевой сенсорной системы с кровью и ликвором. Выделяются опиоидные медиаторы в синаптических окончаниях нейронов противоболевой системы. Аналгезирующий эффект эндорфинов высокий в головном и спинном мозге, эффект энкефалинов в этих структурах средний, эффект динорфинов в головном мозге низкий, в спинном мозге – высокий.
Рис.1. Взаимодействие основных элементов обезболивающей системы первого уровня: ствол мозга – спиной мозг. (светлые кружки – возбуждающие нейроны, черные – тормозящие).
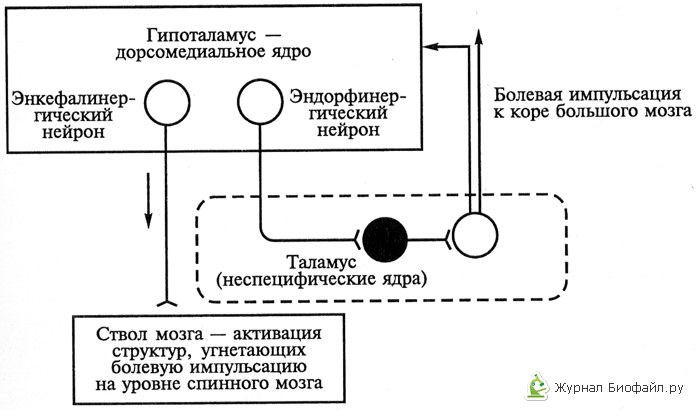
Рис.2. Механизм работы обезболивающей системы организма второго уровня (гипоталамус – таламус – ствол мозга) с помощью опиоидов.
Светлые кружки – возбуждающие нейроны, черные - тормозящие.
Степень выраженности болевого ощущения не определяется одной лишь силой экзогенного или эндогенного болевого воздействия. Во многом оно зависит от соотношения активностей ноцицептивного и антиноцицептивного отделов системы боли, что имеет приспособительное значение.
Поверхностные ткани снабжены нервными окончаниями различных афферентных волокон (Дж.Эрлангер , Г.С.Гассер , 1924). Наиболее толстые, миелинизированные Аb-волокна обладают тактильной чувствительностью. Они возбуждаются при неболезненных прикосновениях и при перемещении. Эти окончания могут служить как полимодальные неспецифические болевые рецепторы только при патологических условиях, например, вследствие возрастания их чувствительности (сенсибилизации) медиаторами воспаления. Слабое раздражение полимодальных неспецифических тактильных рецепторов приводит к чувству зуда . Порог их возбудимости понижают гистамин и серотонин (Г.Штюттген , 1981).
Специфическими первичными болевыми рецепторами (ноцирецепторами) служат два других типа нервных окончаний - тонкие миелинизированные Аd-терминали и тонкие немиелинизированные С-волокна, филогенетически более примитивные. Оба эти типа терминалей представлены и в поверхностных тканях, и во внутренних органах. Некоторые участки тела, например, роговица, иннервируются только Аd и С-афферентами. Ноцирецепторы дают чувство боли в ответ на самые разные интенсивные стимулы - механическое воздействие, термический сигнал (обычно, с температурой более 45-47 0 С), раздражающие химикаты, например, кислоты. Ишемия всегда вызывает боль, поскольку провоцирует ацидоз. Мышечный спазм может вызывать раздражение болевых окончаний из-за относительной гипоксии и ишемии, которые он вызывает, а также вследствие прямого механического смещения ноцирецепторов.
По С-волокнам проводится со скоростью 0,5-2 м/сек медленная, протопатическая, а по миелинизированным, быстропроводящим Аd-волокнам, обеспечивающим скорость проведения от 6 до 30 м/сек, - эпикритическая боль. Кроме кожи, где, по данным А.Г.Бухтиярова (1966), насчитывается не менее 100-200 болевых рецепторов на 1 см 2 , слизистых и роговицы, болевыми рецепторами обоих типов обильно снабжены надкостница (в чём убеждается каждый футболист, получающий при подкате удар по передне-внутренней поверхности голени), а также сосудистые стенки, суставы, мозговые синусы и париетальные листки серозных оболочек.
В висцеральных листках этих оболочек и внутренних органах болевых рецепторов гораздо меньше. К тому же, в паренхиме внутренних органов имеются, исключительно, С-волокна протопатической чувствительности, достигающие спинного мозга в составе вегетативных нервов. Поэтому висцеральную боль труднее локализовать, чем поверхностную. Кроме того, локализация висцеральной боли зависит от феномена “отраженных болей”, механизмы которого рассматриваются ниже. Париетальные брюшина, плевра, перикард, капсулы ретроперитонеальных органов и часть брыжейки имеют не только медленные протопатические С-волокна, но и быстрые эпикритические Аd, связанные со спинным мозгом спинальными нервами. Поэтому боль от их раздражения и повреждения намного острее и чётче локализована. Хирурги еще в доанестезиологическую эпоху заметили, что разрезы кишки менее болезненны, чем рассечение пристеночного листка брюшины. Боли при нейрохирургических операциях максимальны в момент рассечения мозговых оболочек, в то же время кора больших полушарий обладает очень незначительной и строго локальной болевой чувствительностью. Вообще, такой распространённый симптом, как головная боль , практически всегда связан с раздражением болевых рецепторов вне самой ткани мозга. Экстракраниальной причиной головной боли могут быть процессы, локализованные в синусах костей головы, спазм цилиарной и других глазных мышц, тоническое напряжение мышц шеи и скальпа. Интракраниальные причины головной боли - это, в первую очередь, раздражение ноцирецепторов мозговых оболочек. При менингите сильнейшие головные боли охватывают всю голову. Весьма серьёзную головную боль вызывает раздражение ноцирецепторов в мозговых синусах и артериях, особенно в бассейне средне-мозговой артерии. Даже незначительные потери цереброспинальной жидкости (около 20 мл) могут спровоцировать головную боль, особенно, в вертикальном положении тела, поскольку плавчесть мозга меняется, и при уменьшении гидравлической подушки раздражаются болевые рецепторы его оболочек. С другой стороны, избыток цереброспинальной жидкости и нарушение ее оттока при гидроцефалии, отек головного мозга, его набухание при внутриклеточной гипергидратации, полнокровие сосудов мозговых оболочек, вызванное цитокинами при инфекциях, локальные объемные процессы - также провоцируют “самую частую жалобу” - головную боль, так как при этом увеличивается механическое воздействие на болевые рецепторы окружающих собственно мозг структур. Обший принцип локализации головных болей таков, что затылочные боли часто отражают раздражение ноцирецепторов сосудов и мозговых оболочек под tentorium, а надпалаточные раздражители и стимуляция верхней поверхности самой палатки проявляются лобно-теменными болями. Знакомая очень значительной части человечества “головная боль с похмелья” имеет комплексный патогенез, включая индуцированное алкоголем полнокровие мозговых оболочек и внутриклеточную гипергидратацию. Патофизиология некоторых форм головной боли, тесно связанных с гуморальными медиаторами болевой и антиболевой систем и с проводниковыми механизмами этих систем, в частности, мигрени, отдельно рассматривается ниже.
Паренхима селезёнки, почки, печени и легкого совершенно лишена ноцирецепторов. Зато ими богато снабжены бронхи, желчевыводящие пути, капсулы и сосуды этих органов. Даже значительные по размеру абсцессы печени или лёгкого могут быть почти безболезненными. Однако, плеврит или холангит порой дают серьёзный болевой синдром, сами по себе не будучи тяжёлыми. Висцеральные болевые рецепторы отличаются ещё и тем, что развивают сравнительно слабый ответ на строго локальное повреждение органа, например, хирургический разрез. Однако, при диффузном вовлечении ткани в альтерацию (на фоне ишемии, при действии литических ферментов и раздражающих химикатов, при спазмах и перерастяжении полых органов), их чувствительность под воздействием медиаторов воспаления стремительно растёт, и от них исходит сильная импульсация.
Болевые рецепторы претендуют на уникальное положение в человеческом теле. Это единственный тип чувствительных рецепторов, который не подлежит какой бы то ни было адаптации или десенсибилизации под воздействием длящегося или повторяющегося сигнала. Ноцирецепторы не повышают при этом порог своей возбудимости, как это делают другие, например, холодовые сенсоры. Следовательно, рецептор не “привыкает” к боли. Более того, в ноцирецептивных нервных окончаниях имеет место прямо противоположное явление - сенсибилизация болевых рецепторов сигналом. При воспалении, повреждениях тканей (особенно, внутренних органов) и при повторных и длительных болевых раздражителях порог возбудимости ноцирецепторов снижается. Даже легчайшие прикосновения к ожоговой поверхности крайне болезненны. Это явление называется первичной гиперальгезией . Пальпация внутренних органов, даже если она интенсивна, не причиняет боли, если нет их воспаления. Однако при воспалении чувствительность молчащих внутренних ноцирецепторов настолько увеличивается, что врач регистрирует болевые симптомы. Поколачивание по области почек, безболезненное в отсутствие их повреждений, ведет к болевому ощущению в случае, если почечные ноцирецепторы сенсибилизированы медиаторами воспаления (положительный симптом Пастернацкого). Легко отметить, что если бы происходила адаптация болевых рецепторов, все хронические деструктивные процессы были бы безболезненны и боль утратила бы свою функцию сигнала, который, по выражению И.П.Павлова , “побуждает отбросить то, что угрожает жизненному процессу”.
Называя болевые сенсоры рецепторами, мы должны подчеркнуть, что применение к ним этого термина носит условный характер - ведь это свободные нервные окончания, лишённые каких бы то ни было специальных рецепторных приспособлений.
Нейрохимические механизмы раздражения ноцирецепторов хорошо изучены. Их основным стимулятором является брадикинин. В ответ на повреждение клеток близ ноцирецептора освобождаются этот медиатор, а также простагландины, лейкотриены и ионы калия и водорода. Простагландины и лейкотриены сенсибилизируют ноцирецепторы к кининам, а калий и водород облегчают их деполяризацию и возникновение в них электрического афферентного болевого сигнала. Возбуждение распространяется не только афферентно, но и антидромно, в соседние ветви терминали. Там оно приводит к секреции вещества Р. Этот нейропептид, о котором уже упоминалось, вызывает вокруг терминали паракринным путём гиперемию, отек, дегрануляцию тучных клеток и тромбоцитов. Освобождаемые при этом гистамин, серотонин, простагландины сенсибилизируют ноцирецепторы, а химаза и триптаза мастоцитов усиливают продукцию их прямого агониста - брадикинина. Следовательно, при повреждении ноцирецепторы действуют и как сенсоры, и как паракринные провокаторы воспаления. Вблизи ноцирецепторов, как правило, располагаются симпатические норадренергические постганглионарные нервные окончания, которые способны модулировать чувствительность ноцирецепторов. При травмах периферических нервов нередко развивается так называемая каузалгия - патологически повышенная чувствительность ноцирецепторов в области, иннервируемой повреждённым нервом, сопровождаемая жгучими болями и даже признаками воспаления без видимых местных повреждений. Механизм каузалгии связан с гипералгизующим действием симпатических нервов, в частности, выделяемого ими норадреналина, на состояние болевых рецепторов. Возможно, при этом происходит секреция вещества Р и других нейропептидов симпатическими нервами, что и обусловливает воспалительные симптомы. Явление каузалгии представляет собой, в полном смысле, нейрогенное воспаление, хотя оно вызывается не нервным, а паракринным способом (см. также выше, о роли нервной регуляции в воспалении).
Как впервые предположили У.Кэннон и А.Розенблют (1951) паракринная безымпульсная нейропептидэргическая деятельность нервных окончаний в тканях и составляет реальную основу явления, которое в течение более чем 100 лет, от Ф. Мажанди (1824) до Л.А. Орбели (1935) и А.Д. Сперанского , (1937), именовали нервной трофикой .
Дата добавления: 2015-05-19 | Просмотры: 985 | Нарушение авторских прав
| | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | |
В отличие от всех других рецепторов рецепторы болевые рецепторы не имеют какого-либо адекватного раздражителя. Болевые, или ноцицептивные, ощущения могут возникать при действии любых раздражений чрезмерной силы. Так как такие раздражения вызывают повреждения тканей, то возникающие под их влиянием болевые ощущения имеют важное биологическое значение. Они сигнализируют организму об опасности и вызывают оборонительные рефлексы, направленные на устранение раздражения, вызывающего боль. Именно поэтому свыше 200 лет назад французский филосов Вольтер писал, что боль «является верным стражем среди всех наших опасностей; боль громко и непрерывно твердит нам: будьте осторожны, берегите, сохраняйте вашу жизнь».
Болевые ощущения нередко бывают одним из первых, а иногда и единственным проявлением заболевания, позволяющим врачу поставить диагноз, определить тяжесть болезни и необходимые лечебные мероприятия. Однако не всегда имеется соответствие между тяжестью заболевания и интенсивностью ощущений боли. Часто серьезные поражения внутренних органов не сопровождаются болевыми ощущениями и, напротив, нередко сильнейшие болевые ощущения возникают при совершепно нничтожных и не опасных поражениях и являются главной причиной страдания.
Рецепторные аппараты, воспринимающие боль
Вопрос о том, какими нервными структурами воспринимается боль, еще не решен. Одни исследователи считают, что для восприятия боли не существует особых болевых рецепторов , так как чрезмерное раздражение любых рецепторов и нервных стволов способно вызвать чувство боли. Другие считают, что болевые раздражения воспринимаются свободными окончаниями «болевых» нервных волокон.
Главными доказательствами второй точки зрения служат следую факты.
- Существует состояние, называемое анальгезией, при котором отсутствует боль, но сохранено чувство прикосновения (оно возникает при легком наркозе, а также при некоторых заболеваниях спинного мозга) тогда разрез кожи ощущается как прикосновение и давление, но не как боль.
- На коже имеются особые болевые точки: если колоть очень тонкой иглой различные участки кожи, то можно попасть в точки, при уколе которых боль возникает сразу, без предварительного чувства прикосновения. На середине роговой оболочки глаза нет осязательных точек, но имеются болевые; гистологические исследования показали, что там разветвляются только голые веточки чувствительных нервов без всяких специфических осязательных телец.
- После перерезки и сшивания нерва в процессе регенерации нервных волокон сначала восстанавливается болевая чувствительность и лишь затем, через значительное время, остальные виды чувствительности. При восстановлении только болевой чувствительности любое раздражение кожи - прикосновение, поглаживание, давление - вызывает нередко ощущение нестерпимой боли. При восстановлении других видов чувствительности (тактильной, тепловой, холодовой) чрезмерные болевые ощущения исчезают и болевые чувства приобретают обычный характер. Существенно, что такая последовательность восстановления ощущений после повреждения нерва соответствует определенным морфологическим этапам регенерации поврежденных нервных стволов и рецепторов. На ранних этапах регенерации нервных волокон они не имеют миелиновой оболочки и представляют собой свободные нервные окончания (голые осевые цилиндры). Именно в это время любое раздражение воспринимается как боль. По мере появления миелиновой оболочки и восстановления структуры рецепторов возникает и обычная чувствительность кожи, а чрезмерные болевые ощущения исчезают.
Волокна, проводящие болевые импульсы
Электрофизиологические исследования афферентной импульсации нервных стволов и волокон при болевых раздражениях показали, что импульсы, вызывающие ощущение боли, проводятся афферентными волокнами двух типов. Одни из них относятся к группе Аδ, это тонкие миелиновые волокна, со скоростью проведения возбуждения 5-15 м/сек. Другие - тонкие безмиелиновые волокна, относящиеся к группе С, со скоростью проведення возбуждения 1-2 м/сек. Соответственно различной скорости распространения болевых импульсов, а значит и различному времени поступления их в центральную нервную систему, болевые раздражения вызывают как бы двойное ощущение - вначале мимолетное, точно локализованное, но не очень сильное, которое сменяется разлитым «тупым», субъективно весьма неприятным, сильным болевым ощущением.
Существует предположение, что чувство боли возникает в тех случаях, когда появляются синхронные нервные разряды одновременно в очень больном числе афферентных волокон. Это предположение помогает понять тот факт, что при регенерации нервных волокон, когда миелиновая оболочка еще не сформировалась, любое раздражение кожных рецепторов воспринимается как болевое. Отсутствие миелиновой оболочки облегчает вовлекшие в процесс возбуждения одновременпо большого количества нервных волокон.
Адаптация болевых рецепторов
Адаптация болевых рецепторов может быть обнаружена следующим опытом: если в кожу вколоть иглу и не смещать ее, то возникающие от укола нервные импульсы и ощущение боли прекращаются. Они появляются вновь при всяком движении, так как при этом происходит смещение или и раздражение новых неадаптировавшихся болевых рецепторов ( ).
Болевые рефлексы
Болевые раздражения вызывают многообразные рефлекторные реакции. Характерной их особенностью является то, что в осуществление рефлекторного акта вовлекаются многие органы тела.
При болевых рефлексах наблюдаются: повышение мышечного тонуса, учащение сердечной деятельности и дыхания, сужение сосудов, повышение артериального давления, уменьшение мочеотделения и секреции пищеварительных соков, увеличение потоотделения, торможение моторной деятельности кишечника, повышение содержания сахара в крови и увеличенный распад гликогена, сужение зрачков и ряд других явлений. Многие из перечисленных реакций являются следствием возбуждения симпатической нервной системы и усиленной секреции адреналина и гормонов задней доли гипофиза. Увеличена также секреция кортникостероидов. Все перечисленные вегетативные компоненты болевых рефлексов имеют значение в мобилизации сил организма, что необходимо в жизнеопасных ситуациях, когда происходит повреждение тканей, вызывающее болевые ощущения.
Определение локализации болевых раздражений и отраженные боли
Человек хорошо определяет болезненные участки на поверхности кожи. Вместе с тем способность локализовать место болевого раздражения при болях во внутренних органах часто недостаточно отчетливо выражена. При заболеваниях внутренних органов боль может ощущаться в месте заболевания, а в других частях тела, например на поверхности кожи. Такие боли называются отраженными.
Примером могут служить боли во время приступа грудной жабы, т. е. при спазме венечных сосудов сердца, когда возникают болевые ощущении не только в области сердца, но нередко в левой руке и лопатке, в левой половине шеи и головы. Эти отражённые болевые ощущения могут быть гораздо сильнее, чем боли в области сердца. При заболеваниях других внутренних органов также наблюдаются отраженные в определенных участках кожи. Тот кожный участок, в котором возникают 6оли при поражении определенного внутреннего органа, называют зоной Захарьина-Геда.
Ощущения боли, возникающие при раздражении кожи, характеризуются более совершенной локализацией, по-видимому, оттого, что одновременно с болевыми точками кожи раздражаются и тактильные рецепторы, раздражение которых человек точно локализует.
Своеобразным неприятным ощущением, возникающим при раздражении рецептров кожи, является зуд, который вызывает рефлекторную реакцию почесывания кожи. Ощущение зуда связывают с болевыми рецепторами, расположенными под эпидермисом. О роли именно болевых рецепторов свидетельствует тот факт, что потери тактильной чувствительности не сопровождается исчезновением зуда, а потеря болевой чувствительности под влиянием местных обезболивающих (например, кокаина) прекращает зуд.
Рецепторами, при раздражении которых возникает зуд, являются свободные нервные окончания, расположенные под эпидермисом, и связанные с тонкими безмякотными нервными волокнами.
В происхождении зуда имеет значение образование в коже некоторых химических соединений, раздражающих рецепторы. К числу таких веществ некоторые исследователи oтносят гистамин, подкожное введение которого в очень малой дозе вызывает резкий зуд, сопровождающийся расширением капилляров и образованием волдыря. Еще более активны, чем гистамин, некоторые пептидазы-ферменты, расщепляющие полипептиды. При внутрикожном их, введении в ничтожных количествах они вызывают нестерпимый зуд. Действие этих веществ считается специфичным, так как под их влиянием появляется зуд и нет никаких признаков расширения капилляров, воспаления волдыря.
Боль - это величайший эволюционный механизм, который позволяет человеку вовремя замечать опасность и реагировать на нее. Рецепторы болевой чувствительности - это особые клетки, которые отвечают за прием информации, а затем передают ее к мозгу в болевой центр. Подробнее о том, где расположены эти нервные клетки и каким образом они действуют, вы сможете прочесть в этой статье.
Боль
Боль - это неприятное ощущение, которое передают нашему мозгу нейроны. Дискомфорт появляется не просто так: он сигнализирует о фактическом или потенциальном повреждении в организме. Например, если поднести руку слишком близко к огню, здоровый человек тут же ее отдернет. Это мощнейший защитный механизм, который моментально сигнализирует о возможных или текущих неполадках и заставляет нас сделать все, чтобы исправить их. Зачастую боль свидетельствует о конкретных травмах или повреждениях, но также она может носить и хронический, выматывающий характер. У некоторых людей болевые рецепторы обладают повышенной чувствительностью, в результате чего у них появляется боязнь любых прикосновений, так как они вызывают дискомфортные ощущения.
Знать принцип действия ноцицепторов в здоровом организме нужно для того, чтобы понять, с чем связан болевой синдром, как его лечить, а также какие причины вызывают чрезмерную чувствительность нейронов. В настоящее время Всемирная Организация Здравоохранения признала, что ни один человек не должен терпеть боль любого рода. На рынке существует множество препаратов, которые могут полностью купировать или заметно уменьшить болевые ощущения даже у раковых больных.
Зачем нужна боль?

Чаще всего болевые ощущения появляются из-за травмы или болезни. Что происходит в организме, когда, например, мы дотрагиваемся до острого предмета? В это время рецепторы, находящиеся на поверхности нашей кожи, распознают чрезмерную стимуляцию. Мы еще не чувствуем боли, хотя сигнал о ней уже мчится по синапсам к мозгу. Получив сообщение, мозг дает сигнал действовать, и мы отдергиваем руку. Весь этот сложный механизм занимает буквально тысячные доли секунды, ведь от скорости реакции зависит жизнь человека.
Болевые рецепторы на волосяном покрове расположены буквально везде, и это позволяет коже оставаться чрезвычайно чувствительной и чутко реагировать на малейший дискомфорт. Ноцицепторы способны реагировать на интенсивность ощущений, повышение температуры, а также различные химические изменения. Поэтому выражение «боль только в твоей голове» верно, так как именно мозг образовывает неприятные ощущения, заставляющие человека избегать опасности.
Ноцицепторы
Болевой рецептор представляет собой особый тип нервных клеток, которые отвечают за прием и передачу сигналов о различных стимуляциях, которые затем передаются в центральную нервную систему. Рецепторы выпускают химикаты под названием нейротрансмиттеры, которые с огромной скоростью проходят через нервы, спинной мозг к главному «компьютеру» человека в болевой центр. Весь процесс передачи сигналов называется ноцицепцией, а болевые рецепторы, которые расположены в большинстве известных тканей, - ноцицепторами.
Механизм действия ноцицепторов

Как действуют болевые рецепторы в мозге? Они активируются в ответ на какую-либо стимуляцию, будь она внутренней или внешней. В качестве внешней стимуляции можно привести в пример острую булавку, до которой вы случайно дотронулись пальцем. Внутренняя стимуляция может быть вызвана ноцицепторами, расположенными во внутренних органах или костях, например, остеохондрозом или искривлением позвоночника.
Ноцицепторы представляют собой мембранные белки, которые распознают два вида воздействия на мембрану нейрона: физическое и химическое. Когда ткани человека повреждены, рецепторы активируются, что приводит к открытию катионных каналов. В результате, возбуждаются, и в мозг посылается сигнал боли. В зависимости от того, какого рода воздействие оказано на ткани, выделяются разные химические вещества. Мозг обрабатывает их и выбирает «стратегию», по которой нужно действовать. Кроме того, болевые рецепторы не только принимают сигнал и передают его в мозг, но и выделяют биологически активные соединения. Они расширяют сосуды, способствуют привлечению клеток иммунной системы, которые, в свою очередь, помогают быстрее восстановиться организму.
Где они расположены

Человека пронизывает все тело от кончиков пальцев до живота. Она позволяет ощущать и контролировать все тело, отвечает за координацию и передачу сигналов от мозга к различным органам. В этот сложнейший механизм также входит оповещение о травме или каком-либо повреждении, которое начинается с болевых рецепторов. Они располагаются практически во всех нервных окончаниях, хотя наиболее часто их можно встретить в коже, мышцах и суставах. Также они распространены в соединительных тканях и во внутренних органах. На одном квадратном сантиметре кожи человека расположено от 100 до 200 нейронов, которые обладают возможностью реагировать на изменения в окружающей среде. Иногда эта поразительная способность человеческого организма приносит немало проблем, но, в основном, помогает спасти жизнь. Хоть временами нам и хочется избавиться от боли и ничего не чувствовать, эта чувствительность необходима для выживания.
Болевые рецепторы кожи обладают, пожалуй, наибольшим распространением. Однако ноцицепторы можно найти даже в зубах и надкостнице. В здоровом организме любая боль является сигналом о каких-либо неполадках, и ее ни в коем случае нельзя игнорировать.
Различие в типах нерва
Наука, изучающая процесс возникновения боли и ее механизмы, является достаточно сложной для понимания. Однако, если взять за основу знания о нервной системе, то все может оказаться куда проще. Периферическая нервная система является ключевой для организма человека. Она выходит за пределы головного и спинного мозга, поэтому с помощью нее человек не может думать или дышать. Зато она служит отличным «датчиком», который способен уловить мельчайшие изменения как внутри тела, так и снаружи. Состоит она из черепных, спинальных и афферентных нервов. Именно афферентные нервы располагаются в тканях и органах и передают сигнал в мозг об их состоянии. В тканях есть несколько видов афферентных ноцицепторов: А-дельта и С-сенсорные волокна.
Волокна А-дельта покрыты своеобразным гладким защитным экраном, благодаря чему они передают болевые импульсы быстрее всего. Они реагируют на острую и четко локализованную боль, которая требует незамедлительных действий. К такой боли можно отнести ожог, рану, травму и другие повреждения. Чаще всего А-дельта волокна расположены в мягких тканях и в мышцах.

С-сенсорные болевые волокна, напротив, активируются в ответ на неинтенсивные, но длительные болевые стимулы, которые не имеют четкой локализации. Они не миелинизированы (не покрыты гладкой оболочкой) и поэтому передают сигнал в мозг несколько медленнее. Чаще всего эти боевые волокна реагируют на повреждение внутренних органов.
Путешествие сигнала боли
Как только болевой стимул передается по афферентным волокнам, он должен пройти через спинной рог спинного мозга. Это своеобразный ретранслятор, который сортирует сигналы и передает их в соответствующие разделы мозга. Некоторые болевые стимулы передаются непосредственно в таламус или головной мозг, что позволяет дать быстрый ответ в виде действия. Другие отправляются в лобную кору для дальнейшей обработки. Именно в лобной коре возникает сознательная реализация боли, которую мы чувствуем. Из-за этого механизма, во время экстренных ситуаций, мы даже не успеваем почувствовать неприятные ощущения в первые секунды. Например, при ожоге самая сильная боль наступает через несколько минут.
Реакция мозга
Последним шагом в процессе передачи сигнала о боли является ответ от мозга, который сообщает организму, как ему нужно реагировать. Эти импульсы передаются по эфферентным черепным нервам. Во время передачи сигнала о боли в головном и спинном мозге выделяются разнообразные химические соединения, которые либо уменьшают, либо увеличивают восприятие болевого стимула. Они называются нейрохимическими медиаторами. В их состав входят эндорфины, которые являются натуральными анальгетиками, а также серотонин и норадреналин, которые усиливают восприятие боли человеком.
Виды болевых рецепторов

Ноцицепторы делят на несколько видов, каждый из которых является чувствительным лишь к одному виду раздражения.
- Рецепторы температуры и химических раздражителей. Рецептор, отвечающий за восприятие этих стимулов, получил название TRPV1. Его начали изучать еще в 20 веке для того, чтобы получить лекарство, способное избавить от боли. TRPV1 играет роль при онкологии, болезнях дыхательных путей и многих других.
- Пуриновые рецепторы реагируют на повреждение тканей. При этом в межклеточное пространство попадают молекулы АТФ, которые в свою очередь влияют на пуринергические рецепторы, запускающие болевой стимул.
- Рецепторы кислоты. Многие клетки обладают кислоточувствительными ионными каналами, которые могут реагировать на различные химические соединения.
Разнообразие видов болевых рецепторов позволяет быстро передавать в мозг сигнал о наиболее опасных повреждениях и вырабатывать соответствующие химические соединения.
Типы боли
Почему иногда так сильно что-то болит? Как избавиться от боли? Этими вопросами человечество задавалось несколько веков и вот, наконец, нашло ответ. Существует несколько типов боли - острая и хроническая. Острая часто появляется из-за повреждения тканей, например, при переломе кости. Также она может быть связана с головными болями (которыми страдает большая часть человечества). Острая боль уходит так же быстро, как и появляется - как правило сразу после того, как источник боли (например, поврежденный зуб) будет удален.
С хронической болью дело обстоит несколько сложнее. Медики до сих пор не могут полностью избавить своих пациентов от застарелых травм, которые беспокоят их на протяжении многих лет. Хронические боли обычно связаны с длительными болезнями, неустановленными причинами, раком или дегенеративными заболеваниями. Одним из главных сопутствующих факторов хронической боли - неустановленная причина. У пациентов, которые в течение длительного времени испытывают болевые ощущения часто наблюдается депрессия, а болевые рецепторы видоизменяются. Также нарушается химическая реакция организма. Поэтому врачи делают все возможное, чтобы установить источник боли, а если это невозможно, назначают болеутоляющие препараты.
Болеутоляющие препараты
Обезболивающие, или болеутоляющие, препараты, как их иногда называют, обычно работают при помощи нейрохимических медиаторов. Если препарат ингибирует высвобождение «вторичных мессенджеров», то болевые рецепторы просто не активируются, в результате чего сигнал не доходит до мозга. То же самое происходит, если реакция мозга в ответ на раздражитель нейтрализуется. В большинстве случаев обезболивающие могут только временно повлиять на ситуацию, но не могут вылечить основную проблему. Все, что в их силах - это не давать человеку чувствовать боль, связанную с хроническим заболеванием или травмой.

Итоги
Болевые рецепторы волосяного покрова, лимфы и крови позволяют организму человека быстро реагировать на внешние стимулы: изменение температуры, давление, химические кислоты и повреждение тканей. Информация активирует ноцицепторы, которые отправляют сигнал по периферической нервной системе в мозг. Тот, в свою очередь, немедленно реагирует и посылает обратный импульс. В результате мы отдергиваем руку от огня раньше, чем успеваем осознать это, что позволяет существенно снизить степень повреждений. Болевые рецепторы имеют, пожалуй, такое влияние на нас в экстренных ситуациях.
|
Ярослав Алексеевич Андреев - кандидат биологических наук, старший научный сотрудник лаборатории нейрорецепторов и нейрорегуляторов отдела молекулярной нейробиологии Института биоорганической химии им. академиков М. М. Шемякина и Ю. А. Овчинникова РАН. Научные интересы связаны с поиском и характеристикой модуляторов болевых рецепторов. |
|
Юлия Александровна Логашина - младший научный сотрудник той же лаборатории. Занимается поиском и характеристикой новых лигандов TRPA1 рецептора. |
|
Ксения Игоревна Лубова - студентка биологического факультета Московского государственного университета им. М. В. Ломоносова. Изучает TRP рецепторы и их модуляторы. |
|
Александр Александрович Василевский - кандидат химических наук, руководитель группы молекулярных инструментов для нейробиологии отдела молекулярной нейробиологии Института биоорганической химии им. академиков М. М. Шемякина и Ю. А. Овчинникова РАН. Специалист в области ионных каналов и природных токсинов. |
|
Сергей Александрович Козлов - доктор химических наук, руководитель лаборатории нейрорецепторов и нейрорегуляторов того же отдела. Область научных интересов - белковые рецепторы в нервной системе и их лиганды. |
Говорят, что жизнь - это боль. Хотя в этой фразе содержится нечто негативное, связанное с неприятными ощущениями, переживаниями или даже тяжелыми страданиями, не стоит забывать, что боль (ноцицепция) предупреждает нас об опасности - сигнализирует о нарушениях в организме, который немедленно принимается их устранять. Вместе с тем существует и боль, которая приносит только мучения.
Основная причина появления такой боли - сбои в передаче болевых сигналов (нервных импульсов) от чувствительных нейронов к головному мозгу, который и формирует неприятные ощущения. Когда воздействие неопасных стимулов распознающие нейроны расценивают как опасное, развивается состояние, которое называется гиперчувствительностью. И это не всегда плохо, так как в нужный момент она играет важную роль в процессе выздоровления и восстановления организма. Однако бывает и так, что реального повода нет, а гиперчувствительность ведет к изнурительной хронической боли. В таком случае самые обычные безобидные стимулы (легкое прикосновение или тепло) вызывают аллодинию (от греч. άλλος - другой и οδύνη - мучение), а болезненные стимулы - боль еще большей интенсивности, гиперальгезию (от греч. ὑπέρ - сверх- и ἄλγος - боль). Часто аномально интенсивная и нередко хроническая боль, которая изматывает и физиологически, и психологически, а также затрудняет выздоровление, возникает в результате таких заболеваний, как артрит, опоясывающий лишай, СПИД, рак костей и др.
Прежде чем винить в аномалиях чувствительные нейроны (ноцицепторы), которые воспринимают, анализируют и передают болевые сигналы, разберемся, как они работают в здоровом организме и что происходит при патологиях.
Почему так больно?
Биологическая функция ноцицепторов состоит не только в регистрации раздражителя и сообщении об этом нашему мозгу, но и в восприятии сигналов от ближайших соседей. Нейроны окружены другими клетками организма и межклеточной средой, за сохранность и правильное функционирование которых отвечает наша нервная система. Поэтому у ноцицепторов имеется множество молекулярных сенсоров (или рецепторов), настроенных на распознавание химических раздражителей, изменения состава и свойств межклеточной среды, выброса сигнальных молекул из близлежащих клеток. Нейрон самостоятельно «вычисляет» вклад каждого такого молекулярного сенсора по силе и длительности стимуляции, и, если стимулы расцениваются как нежелательные, сигнализирует об этом - и нам становится больно; это «нормальная» физиологическая боль (ноцицепция). Патологическая боль возникает как в случае гибели нейронов при повреждении проводящей сети периферической или центральной нервной системы, так и при ошибочной работе самих нейронов, а ошибаются они из-за неправильной работы их сенсоров.
Болевые сенсоры (или рецепторы) - это мембранные белки, которые распознают физическое или химическое воздействие на мембрану нейрона. При этом они являются катион-селективными ионными каналами, то есть обеспечивают проведение положительно заряженных ионов (натрия, калия, кальция) через клеточную мембрану. Активация рецепторов приводит к открытию катионных каналов и возбуждению чувствительных нейронов - возникновению нервного импульса. Подробнее о наиболее изученных болевых рецепторах мы расскажем ниже.
Что происходит, когда, предположим, человек по неосторожности обжег руку горячим предметом? Такое опасное температурное воздействие регистрируют рецепторы, которые располагаются в мембране ноцицептора. Они мгновенно распознают сильную стимуляцию и передают импульс в центральную нервную систему. На столь сильное возбуждение мозг незамедлительно реагирует, и мы рефлекторно отдергиваем руку от горячего предмета. Интересно, что те же сенсоры реагируют на капсаицин - активное вещество жгучего перца, вызывающего «пожар» во рту.
За распознавание ряда опасных химических воздействий отвечают другие рецепторы, которые воспринимают стимулы только с внутриклеточной стороны, поэтому для их активации опасные вещества должны не только проникнуть через кожу, но и попасть внутрь нейрона, «пробравшись» через липидную биомембрану. Если химический ожог вызван кислотой, то работать будет именно тот рецептор, который чувствителен к изменению кислотности среды, и тоже даст сильный ответ, как только кислота достигнет нейрона.
Руку мы отдернули, но за время контакта с горячей поверхностью часть наших клеток погибла, и в ответ на повреждение ткани у нас начинает развиваться воспалительный процесс. В этом тоже принимает участие наша нервная система. Из поврежденных клеток через разорванные цитоплазматические мембраны во внеклеточную среду начинают выделяться характерные для внутриклеточной среды молекулы, в частности аденозинтрифосфорной кислоты (АТФ). На этот случай в нейронах тоже есть свой рецептор, который активируется молекулами АТФ и сигнализирует о том, что рядом с ним произошла гибель клеток и требуется их восстановление. Дело в том, что АТФ, как известно еще со школы, - основная энергетическая молекула организма, и такая «ценность» редко оказывается в межклеточной среде.
Нейрон не просто сигнализирует, он выбрасывает во внеклеточную среду особые биологически активные соединения, медиаторы воспаления, что приводит к длительному развитию нейрогенного воспаления - расширению сосудов и привлечению клеток иммунной системы. Пока идет процесс регенерации и в среде присутствуют медиаторы воспаления, сенсорные нейроны посылают сигнал в центральную нервную систему, где он тоже воспринимается как боль, но уже не такая сильная. Так как поврежденная ткань нуждается в защите, чувствительность нейронов к внешним воздействиям повышается, и даже незначительное механическое или тепловое воздействие будет вызывать сильную болевую реакцию. Это и есть «полезная» гиперчувствительность.
Почти все знают, что к поврежденной ткани рекомендуется приложить холод, чтобы облегчить боль и уменьшить воспаление. В этом эффекте также задействованы нейрональные рецепторы. Главный рецептор «по холоду» - ментоловый (помните «мятный» холодок?) - находится не в тех же нейронах, где располагается «тепловой», а потому ощущения холода и жары передаются различными чувствительными волокнами. Оказывается, информация от разных ноцицепторов «суммируется» в спинном мозгу, сигнал от горячего воздействия корректируется с учетом сигнала от холодного, и именно поэтому приложенный кусочек льда может унять сильную боль.
Описанная схема развития боли сильно упрощена (рис. 1). На самом деле, чтобы разобраться в деталях ноцицепции, ученые исследуют каждый рецептор отдельно в изолированных условиях. Эксперименты проводят на клеточных линиях, в которые методами генной инженерии встраивают гены определенных рецепторов. Расскажем немного об изучении и функциях нескольких наиболее важных болевых рецепторов. Как оказалось, они не всегда ориентированы на распознавание и генерацию болевого сигнала, но вовлечены в регуляцию многих других процессов, поэтому умение корректировать их работу различными лекарственными препаратами поможет лечить разнообразные болезни (рис. 2).

Рецепторы температуры и химических раздражителей
Очень часто в развитии боли и воспаления играют роль чувствительные нейроны, которые отвечают за восприятие высокой температуры. Еще в середине XX века обнаружили, что большие дозы капсаицина вызывают у экспериментальных животных новый тип обезболивания (анальгезии) . После введения капсаицина вначале наблюдается характерная поведенческая реакция, вызванная болью, но затем наступает длительный период потери чувствительности к ряду внешних стимулов. Животные в таком состоянии нормально реагируют на мягкое механическое раздражение, но утрачивают реакцию на многие болевые стимулы, и у них не развивается нейрогенное воспаление. Таким образом, нейроны, отвечающие за восприятие высокой температуры, также отвечают за восприятие химических раздражителей и нейрогенный компонент воспалительного ответа . Стало очевидно, что рецептор, который реагирует на воздействие температуры и капсаицина, может оказаться полезной мишенью для поиска средств, направленных на лечение воспаления и боли . В конце ХХ в. этот рецептор был охарактеризован на молекулярном уровне и назван TRPV1 (от англ. transient receptor potential channel vanilloid family member 1 - первый представитель ванилоидного семейства рецепторов переменного рецепторного потенциала), или проще - ванилоидный рецептор 1 (рис. 3) . Название «ванилоидные рецепторы» дано не случайно: TRPV1 и другие представители семейства активируются химическими соединениями, содержащими ванилиновую группу (например, капсаицином). Установлено, что TRPV1 - катион-селективный ионный канал, который активируется различными стимулами (температурой выше 43°C, низким рН, капсаицином), а кроме того, его активность регулируется медиаторами воспаления, правда, не напрямую, а через внутриклеточных посредников. Мыши, нокаутные по гену TRPV1 (то есть такие, у которых ген этого рецептора отсутствует или поврежден так, что не работает), значительно медленнее реагируют на тепло, и у них почти не появляется тепловая гиперчувствительность при воспалении . TRPV1 играет важную роль в ряде патологических состояний: при болях, вызванных воспалительным процессом, при онкологических, нейропатических и висцеральных болях, а также при заболеваниях дыхательных путей, панкреатите и мигрени.

Исследования TRPV1 привели к интенсивному изучению подобных рецепторов. Так, был обнаружен еще один ванилоидный рецептор - TRPV3. Интересно, что он реагирует как на приятное тепло, так и на болезненный жар: активность TRPV3 регистрируется при температуре выше 33°C, причем его ответ сильнее на более высокую температуру и возрастает при повторяющейся тепловой стимуляции. Помимо температуры, этот рецептор также активируется камфорой, едкими экстрактами тимьяна, орегано и гвоздики. TRPV3 - еще один кандидат на роль участника в болевой гиперчувствительности, его активность регулируется медиаторами воспаления. Наконец, он напрямую активируется оксидом азота II (NO) - вторичным мессенджером, обеспечивающим увеличение чувствительности нейронов к стимуляции. Также следует отметить наличие TRPV3 в клетках кожи кератиноцитах, где его активация приводит к выбросу воспалительного медиатора интерлейкина-1, что подчеркивает важную роль этого рецептора в воспалительных заболеваниях кожи .
TRP-рецепторы - тетрамеры (рис. 3), то есть образованы четырьмя полипептидными цепочками. При этом могут собираться как гомомеры, то есть рецепторы, сформированные одинаковыми цепочками (например, TRPV1 или TRPV3, описанные выше), так и гетеромеры из разных цепей. Гетеромерные рецепторы (например, построенные из цепочек TRPV1 и TRPV3) обладают различной чувствительностью к тепловым стимулам, пороговая температура их активации лежит между значениями, пороговыми для гомомерных рецепторов.
Интересна история открытия холодового рецептора TRPM8 (здесь «M» означает «меластатин», что указывает на функцию рецепторов этого семейства в меланоцитах - клетках кожи, ответственных за пигментацию). Вначале был обнаружен кодирующий его ген, активность которого повышалась при раке простаты и некоторых других онкологических заболеваниях . Много позже была показана способность TRPM8 реагировать на ментол (компонент мяты) и ряд других «освежающих» веществ, а также на понижение температуры (ниже 26°С). Теперь этот рецептор считается основным сенсором холода в нервной системе . Исследования выявили, что TRPM8 отвечает за широкий диапазон восприятия холодовых стимулов - от приятной прохлады до болезненного холода и холодовой гиперчувствительности. Такое разнообразие функций объясняется существованием нескольких субпопуляций чувствительных нейронов, которые используют TRPM8 как многофункциональный сенсор холода, настроенный на определенную температуру при участии внутриклеточных сигнальных систем.
Самый непонятный и очень важный рецептор TRPA1 (здесь «A» означает «анкирин», что указывает на наличие в структуре рецепторов этого семейства большого числа «анкириновых повторов», особых белковых элементов) находят в чувствительных нейронах кожи, клетках эпителия кишечника, легких и мочевого пузыря, причем TRPA1 часто соседствует с TRPV1 . Вещества, активирующие TRPA1, вызывают жжение, механическую и термическую гиперчувствительность, а также нейрогенное воспаление. Гиперэкспрессия гена, кодирующего TRPA1, ведет к возникновению хронического кожного зуда и аллергического дерматита. Наследственное заболевание «синдром эпизодической боли», которое характеризуется неожиданно возникающей изнурительной болью при голодании или физической нагрузке, связано с мутацией в этом рецепторе, приводящей к его избыточной активности .
Основная функция TRPA1 - распознавание химических и воспалительных агентов, и их ассортимент столь велик, что с правильной работой этого рецептора связаны почти все жизненные процессы нашего организма. В дыхательной системе он распознает летучие вредные вещества: слезоточивый газ, озон, альдегиды (акролеин, компоненты корицы), сераорганические соединения (жгучие компоненты горчицы, лука и чеснока), вызывая кашель, чихание и образование слизи. В кишечнике TRPA1 регистрирует присутствие воспалительных агентов. Гиперактивность мочевого пузыря при диабете вызвана активацией этого рецептора акролеином, который накапливается в моче. Выявлено участие TRPA1 в возникновении мигрени под влиянием сигаретного дыма и формальдегида у некоторых людей .
Воздействие на рецепторы чувствительных нейронов, участвующие в восприятии температуры, с помощью лекарственных средств приводит к облегчению боли и воспаления. Именно так, не зная о молекулярных мишенях, народная медицина в разное время применяла настойки перца (TRPV1), горчицы (TRPA1), мяты (TRPM8) и гвоздики (TRPV3) для лечения ряда воспалительных заболеваний.
Пуриновые рецепторы
Мы уже упоминали, что организму очень важно знать о повреждении тканей. При травмах, когда нарушается целостность органов и происходит гибель клеток, при ишемии или воспалении в межклеточное пространство попадают молекулы АТФ. Этот кофермент множества реакций обеспечивает энергией многие процессы в клетке; он слишком ценен для функционирования клеток, поэтому редко выбрасывается за их пределы. Восприятие повышения локальной концентрации АТФ осуществляют пуринергические рецепторы (P2X), являющиеся катион-селективными ионными каналами, они запускают болевой ответ, возникающий вследствие разрушения тканей, деформации органов и развития опухолей . Для чувствительных нейронов характерны подтипы P2X2 и P2X3, важная роль последнего в развитии боли при воспалении показана в исследованиях на нокаутных мышах. Также известно, что P2X-рецепторы имеют принципиальное значение для многих физиологических процессов, таких как регуляция тонуса сосудов, вкусовая рецепция и т.д.
Рецепторы кислоты
Для регистрации кислотности во многих типах клеток нервной системы присутствуют так называемые кислоточувствительные ионные каналы (acid-sensing ion channels , ASIC). Считается, что они осуществляют передачу сигнала, связанного с локальным изменением рН при нормальной нейрональной активности в центральной нервной системе. Однако задействованы они и в патологических процессах. В последнее время рецептор подтипа ASIC1a рассматривается как один из основных факторов гибели нейронов в центральной нервной системе при ишемических состояниях. При ишемии и гипоксии усиливается гликолиз, в результате чего происходит накопление молочной кислоты и последующее «закисление» ткани. «Выключение» рецептора ASIC1a вызывает нейропротекторное действие в модели ишемии, что было показано на нокаутных мышах . В периферической нервной системе и тканях внутренних органов ASIC ответственны за чувствительность к боли, возникающей при тканевом ацидозе в мышцах, при сердечной ишемии, повреждении роговицы, воспалении, новообразованиях и местной инфекции . В нейронах периферической нервной системы в основном представлены рецепторы подтипа ASIC3, активность которых также необходимо понижать для купирования боли.
В отличие от TRP-рецепторов, P2X-рецепторы и ASIC являются тримерами (рис. 3), т.е. собраны из трех полипептидных цепочек. Но точно так же эти рецепторы могут быть гомомерами и гетеромерами, что увеличивает их разнообразие и спектр выполняемых функций.
Как победить боль?
Так что же делать, если мы испытываем боль? Если это боль острая или хроническая, терпеть ее нельзя, и необходимо использовать обезболивающие средства, чтобы вернуть нашу систему ноцицепции в нормальное состояние, а себя - к жизни в самом прямом смысле этого слова. В настоящее время для обезболивания применяется множество лекарственных препаратов различных фармакологических групп. Основное место в этом ряду занимают нестероидные противовоспалительные средства (НПВС), антиконвульсанты и антидепрессанты, а также наркотические анальгетики (морфин и другие опиаты и опиоиды). Имеющиеся в настоящее время анальгетические средства влияют главным образом на пути передачи и распространения боли. Для специфичного регулирования рецепторов боли, описанных выше, пока препаратов на рынке лекарств нет.
Первой «болевой» мишенью для фармацевтических компаний стал рецептор TRPV1, поскольку содержащие его чувствительные нейроны играют роль интеграторов многих стимулов, воспринимающихся как боль. Скрининг химических библиотек и рациональный дизайн лигандов на основе знаний о сайте связывания капсаицина позволили создать значительное количество высокоэффективных низкомолекулярных ингибиторов TRPV1. Эти соединения обладали обезболивающим эффектом, но приводили к развитию гипертермии - повышению температуры тела (на 1,5–3°С). Гипертермия стала основной причиной отказа фармакологических компаний от развития лекарственных препаратов на основе полных антагонистов рецептора TRPV1. Однако если ингибировать этот рецептор лишь частично, повышения температуры тела можно избежать. И такие частичные ингибиторы TRPV1 нам, под руководством академика Е. В. Гришина (1946–2016), удалось найти в яде морской анемоны Heteractis crispa . В яде анемоны обнаружено сразу три пептида, ингибирующих TRPV1 и не повышающих температуру тела [ , ], но наиболее мягким эффектом обладал пептид, получивший название АРНС3. Он имеет сильный анальгетический эффект в дозах 0,01–0,1 мг/кг массы тела и слабо понижает температуру тела (всего на 0,6°С) . По силе обезболивания он сопоставим с морфином, но не вызывает наркотического действия и привыкания. По данным доклинических исследований, пептид полностью пригоден для дальнейших клинических испытаний, так как никакие побочные эффекты на лабораторных животных не были обнаружены. Более того, понижение температуры тела необходимо, например, для обеспечения нейропротекции у выживших после остановки сердца, и гипотермическое действие пептида может служить дополнительным бонусом.
Работая под руководством Гришина, мы также обнаружили ингибитор P2X3-рецепторов. Это тоже оказался пептид, которому было дано имя PT1, а найден он был в яде паука Alopecosa marikovskyi . Кстати, PT1 уже успешно прошел лабораторные и доклинические испытания, так что через какое-то время он вполне может стать одним из первых принципиально новых анальгетиков, специфично ингибирующих «болевые» рецепторы. Для третьего из упомянутых подобных рецепторов, ASIC3, нами также был найден ингибитор: пептид Ugr 9-1; источником стал яд морской анемоны Urticina grebelnyi .
Заметим, что в природных ядах часто находят токсины с обратным эффектом, то есть вещества, активирующие рецепторы боли. С точки зрения биологии ядовитых животных это понятно: «болевые» токсины используются ими в целях защиты. Например, в яде китайского птицееда Haplopelma schmidti содержится сильнейший активатор TRPV1, а из яда техасской коралловой змеи Micrurus tener получен активатор ASIC1a. Сейчас уже научились извлекать пользу из таких веществ: их применяют как молекулярные инструменты, чтобы «замораживать» болевые рецепторы в активированном состоянии и исследовать их структуру (рис. 3) [ , ]. С другой стороны, обнаружение полезных молекул в природных ядах - тоже довольно распространенное явление, и несколько природных токсинов (или веществ, созданных на их основе) сегодня применяются в медицине как лекарства. Вот где обретает особый смысл известное изречение средневекового алхимика Парацельса: «Все есть яд, и ничто не лишено ядовитости; одна лишь доза делает яд незаметным».
Рецепторы чувствительных нейронов представляют собой заманчивую, но сложную мишень для создания лекарств. Препараты, если они обладают хорошей селективностью к этим рецепторам, будут приняты потребителями с большой радостью, так как почти все современные средства ограничены в применении из-за побочных эффектов. Работы по поиску селективных препаратов ведутся, в том числе и в нашей стране, и при благоприятном стечении обстоятельств такие лекарства уже скоро смогут появиться в аптеках. Долгих вам лет жизни без боли!
Работа выполнена при поддержке Российского научного фонда (проект № 14-24-00118).
Литература
.
Palermo N. N., Brown H. K., Smith D. L. Selective neurotoxic action of capsaicin on glomerular C-type terminals in rat substantia gelatinosa // Brain Res.
1981. V. 208. P. 506–510.
.
O’Neill J., Brock C., Olesen A. E. et al.